世界各地の110系統の大麻草を全ゲノム解析した国際的な研究
大麻草(Cannabis sativa L.)は、人類が最初に栽培した植物の一つとして、人類社会の経済的社会的、文化的発展に織り込まれた長い変動のある歴史を持っています。
原産地は、中央アジアと言われていますが、野生から栽培化された時期、繊維型と薬用型が分岐した時期については、よくわかっていませんでした。そこで、スイス、中国、英国、カタール、パキスタン、インドの国際研究チームは、世界各地から110系統の大麻草の全ゲノムを対象として研究を行いました(原文は注1を参照)。
集められた系統をゲノム解析すると、大麻草は以下の4つのグループに分類できたのです。
・基底型(中国で採取された14の野生/在来種、米国の2つの野生種)
・繊維型(世界各地のヘンプ系統、野生5種、在来13種、栽培20種)
・薬用型野生(中国南部で採取された3つの野生種、ヒマラヤ以南のインドとパキスタンで採取された11の野生種、およびインドの1品種)
・薬用型(世界各地で栽培されている35種)
原文では、野生(feral)については、野生化した植物、野生にもどった植物と定義されており、単なる野生(wild)と区別されており、純粋な野生の大麻草は絶滅しており、現存していないとしています。
東アジアで12000年前に栽培化の起源
この研究では、コアレセント理論(coalescent theory)と呼ばれる比較的新しい集団遺伝学の理論に基づく、PSMC(pairwise sequentially Markovian coalescent)法を用いて解析をしています。
コアレセント(coalescent)という語は“合着”あるいは“合祖”と訳され、集団のある2つのDNA配列が世代をさかのぼり共通の祖先にいたることをさします。試料として得られたDNA配列の合着が、いつどれだけ起こったかを調べれば、過去の集団における個体数の変動などを推定できるのです。(詳しくは、「注2」を参照)
解析の結果は、祖先種である基底型の大麻草から栽培化が始まったのは、紀元前12,000年頃(95%信頼区間:6458~15,728年)であり、その場所は、東アジア(中国遼寧省付近のNEB;繊維型の在来種)と推定されました。
これは、中国南部や台湾の紐を押した土器の年代(紀元前12,000年)や、日本の縄文時代の遺跡から出土した種子の年代(紀元前10,000年)と一致しています。
東アジアは、イネ、ホウレンソウ、アワ、ダイズ、キクイモ、アプリコット、モモなど、いくつかの作物種が栽培化された、古代の重要な多発地帯(ホットスポット)として知られています。今回の研究により、大麻草もそれらの作物と同様、東アジアで栽培化されたものと思われます。
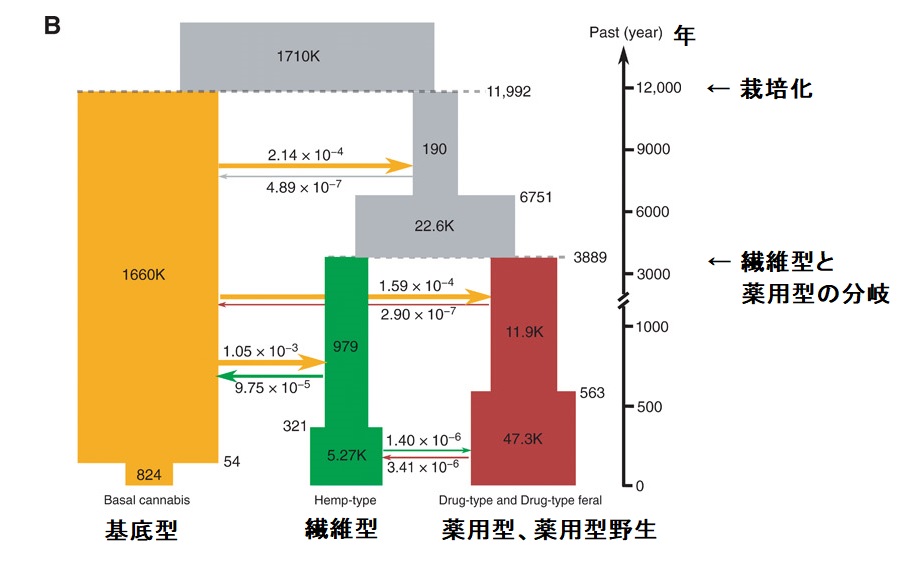
図:栽培化時期と繊維型と薬用型の分岐時期
繊維型ヘンプと薬用型マリファナに分岐したのは4000年前
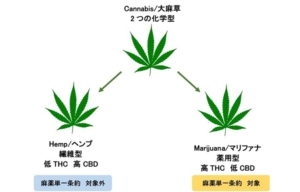
繊維型のヘンプと、薬用型のマリファナのもっとも大きな違いは、精神活性作用のあるTHC濃度です。米国やカナダでは、ヘンプは、THCが0.3%以下の品種であり、それ以上のものは、マリファナ(最近ではカンナビスと呼ぶ場合が多い)と呼ばれています。
大麻草の植物体内では、THCは、THCA(テトラヒドロカンナビノール酸)、CBDはCBDA(カンナビジオール酸)となっており、熱、酸素、光などの影響で脱炭酸化すると、THCやCBDとなります。
遺伝子レベルで見ると、ヘンプは、THCA合成酵素遺伝子が少ないか若しくは欠損し、CBDA合成酵素遺伝子が多く発現している一方で、マリファナは、THCA合成酵素遺伝子が多く、CBDA合成酵素遺伝子が少なく発現していることが明らかとなっています。これらは、栽培化により品種改良の過程においてヘンプは、THCA合成酵素遺伝子を、マリファナはCBDA合成酵素遺伝子を失ってきたことも明らかにしています。
解析の結果、繊維型と薬用型が分岐したのは、紀元前4000年頃でした。中国西部では、少なくとも紀元前2500年頃の考古学的遺跡から大麻草の儀式的な使用や酩酊状態での使用が記録されています。インドにおける大麻草の最初の考古学的な記録は紀元前3000年頃にさかのぼり、この種は他の作物とともに中国から導入されたと考えられています。東アジアとは対照的に、紀元前2000年頃のインドの歴史書には、この種が薬用にのみ利用されていたことが示されています。
この研究では、考古学的な記録と、ゲノム解析の結果はほぼ一致していると結論づけられています。さらに、繊維型と薬用型の初期の栽培化後の分岐に関与する下記のような候補遺伝子を同定しています。
|
繊維型 ヘンプ |
薬用型 マリファナ |
|
枝の形成抑制する遺伝子 (D14、KNAT1) 開花期や光周期に関連する遺伝子 (FLK、EHD3) セルロースやリグニンの生合成に関連する遺伝子(SS、SPS1) |
枝の形成を促進する遺伝子 (NDL2、DTX48) 開花期に関連する遺伝子 (HUA2、FPF1) リグニン生合成に関連する遺伝子 (CSE、C4H) カンナビノイド生合成に必要な基質を生成する酵素をコードする遺伝子(HDR) |
ヘンプの品種選抜と改良は、繊維を大量に採るために、背丈を高くし、枝分かれしないように栽培し、靱皮繊維のセルロース分が多く、リグニンを少なくしてきたことがわかります。
また、マリファナの品種選抜と改良は、花をたくさん産出するために、枝分かれさせ、リグニン豊富な木質の芯をもち、カンナビノイドを大量に生産するようにしてきたことがわかります。
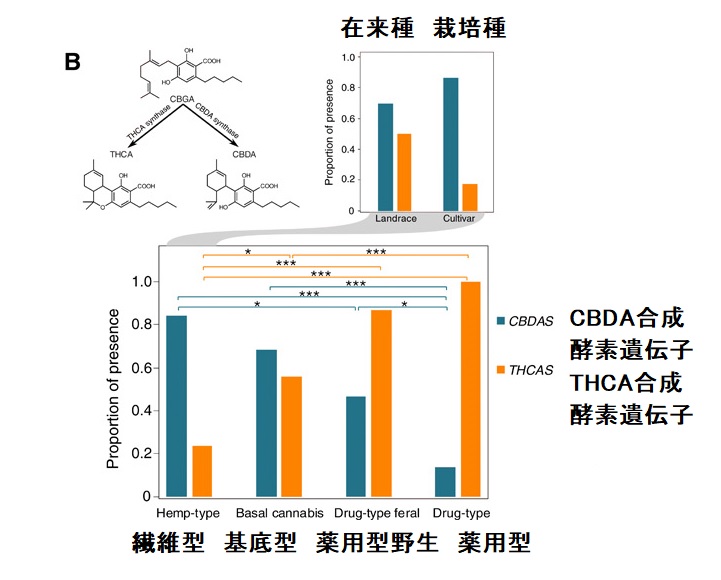
図:各タイプの大麻草のCBDA及びTHCA合成酵素遺伝子の発現比較
今回の研究のまとめ
この研究は、構造的・生化学的に異なる製品を持つ植物の栽培化と世界的な広がりについて、新たな知見を提供した画期的な成果です。
今回の結果は、カンナビノイドの組成が大麻草によって異なるのは、初期の農家がTHCA合成酵素などの機能喪失型の突然変異を人為的に選択した結果であるという進化のシナリオを裏付けるものです。
そして、現在、世界中で進行中の医療や農業の分野で行われている分子育種や機能性研究のための、貴重なゲノム資源の基盤となるでしょう。
日本における大麻ゲノム研究の可能性
今回の国際的な大麻のゲノム研究チームに日本は参加していませんし、日本の麻も解析対象には入っていません。長い歴史を誇る伝統的な麻文化を有し、世界に先駆けて無毒アサの開発に成功した日本として、これはたいへん残念なことです。
実は、本州の栃木県で1983年に開発された無毒アサ品種「とちぎしろ」は、当時の九州大学薬学部正山教授によって佐賀県の在来種から偶然発見されたマリファナの主成分THCが0%の変異種(CBDA種)と栃木県在来種の栃木試1号を両親とする交雑育種法によって育成されたものです。
新しい品種をつくる「育種」の世界では、遺伝資源としての変異種や在来種の大麻は貴重な存在であり、それらの収集・保存・研究がたいへん重要です。新たな品種の開発はおろか、海外の優れたヘンプ品種の輸入すら認めない我が国の大麻政策の後進性によって、日本の大麻研究は世界に大きな遅れを取ってしまいました。
北海道内の野生大麻(自生大麻)は、明治時代に栃木県をはじめとした本州各地から導入された品種が雑草化したものであることがわかっています。これらは、第二次世界大戦後70年以上、人間の手を借りずに北海道の厳しい環境の中で自生してきたことから、生命力が強く、大きな可能性を秘めた世界的に見ても貴重な遺伝資源ですが、これを収集保存するどころか、見つけ次第、駆除し、焼却しているのが現状です。
日本の在来種や北海道の野生大麻のゲノム解析研究は、学問的にも実用的にも貴重な知見をもたらすものと思われます。海外のこうしたゲノム研究を大いに参考にして、近い将来、日本でも新品種育成に向けた農学分野におけるゲノム解析研究が可能となるよう強く希望します。
日本の大麻ゲノム研究を進めるために、資金的&人的な支援をしていただける方はぜひ北海道ヘンプ協会にご連絡をお願いします。
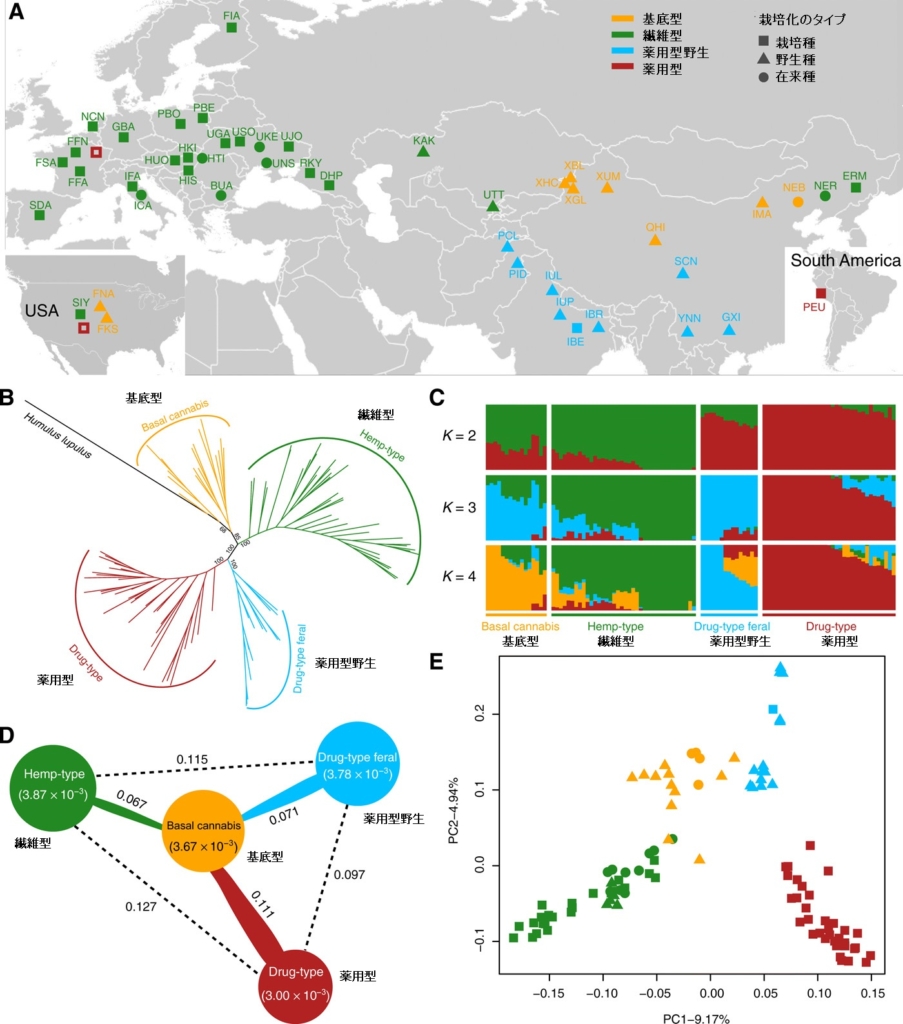
図:集められたゲノム資源の分布地図と基底型、繊維型、薬用型野生、薬用型の系統樹など
注1:原文
Large-scale whole-genome resequencing unravels the domestication history of Cannabis sativa, Science Advances, 7(29) 16 Jul 2021:
https://advances.sciencemag.org/content/7/29/eabg2286
注2:集団ゲノミクスからさぐる植物の進化